



Эволюция живых существ может быть понята только в контексте геологического времени.








Эволюция живых существ может быть понята только в контексте геологического времени.

К концу мела, 65 млн. лет назад, абсолютно все группы динозавров вымерли. Вместе с ними исчезли мозазавры, плезиозавры, птерозавры и целый ряд других наземных и морских животных, в том числе аммониты и белемниты. Вымерло 16% семейств морских животных и 18% семейств наземных позвоночных.

В статье рассказывается о наиболее интересных фактах из жизни древних динозавров. Они были самыми крупными из когда-либо живших на суше существ. В мезозойскую эру эти гигантские ящеры безраздельно властвовали на всей Земле. Затем они полностью вымерли, а по земле распространились наши предки — млекопитающие. Но палеонтологи до сих пор продолжают раскапывать все новые и новые окаменелости этих знаменитых ящеров.

Растения и животные произошли от одного примитивного предка в ходе длительной эволюции. Случилось это несколько миллиардов лет назад.
Биогеография изучает биосферу и географическое распределение организмов и их сообществ (биоценозов) в зависимости от условий обитания и геологических особенностей местности, а также характер фауны и флоры отдельных территорий. Биогеография тесно взаимодействует с биологией и экологией. Составными частями в нее входят зоогеография и фитогеография, предметом изучения которых являются соответственно животные и растения.

Батиметрия занимается изучением рельефа подводных частей морей и океанов. Палеобатиметрия изучает подводные глубины, существовавшие в прошлые геологические эпохи. Условия жизни организмов в морской среде позволяют разделить подводные части океанов в вертикальном направлении на области, различающиеся глубиной и, соответственно, давлением и освещенностью.
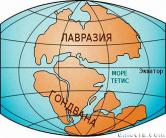
Историческая геология, используя принципы и методы геологии, изучает историю и законы развития Земли со времен образования земной коры в гадейскую эру до нашего времени. Задачи исторической геологии — восстановление хода эволюции земной поверхности и населяющего ее органического мира, а также выяснение истории изменений структуры земной коры.

Бактерии постепенно осваивают кислородный фотосинтез, хотя существенные результаты это даст только в следующую, протерозойскую эру. В конце неоархея находки следов жизни становятся все более распространенными. Среди них стоит отметить группу Малмани, плато Гаап, формации Науга, Когелбеен, Гамохаан и другие.

В неоархее бактериальная жизнь продолжает господствовать в древних земных океанах. Наиболее известные местонахождения следов жизни — это формация Джирина, окрестности огромного стратовулкана Бэк Ривер в Канаде, формация Чешир, субгруппа Шмидтсдриф и другие.

Несмотря на то, что в неоархее жизнь еще не выходила за пределы бактериальных форм, количество находок микрофоссилий и строматолитов постоянно увеличивается. Это такие известные местонахождения, как формация Тумбиана, Маддина, Манджери и находки кратона Дхарвар в Южной Индии.
Кроме этих механизмов сохранения или увеличения существующего числа хромосом, у организмов, очевидно, должны существовать механизмы для его уменьшения. Механизмы уменьшения большого числа хромосом, наблюдающегося при эндополиплоидии, мало изучены, но есть данные о существовании таких механизмов у насекомых и у растений. У огромного большинства организмов имеется общий механизм уменьшения зиготического числа хромосом до гаме - тического; этот механизм называется Мейозом .

После того как хромосомы разошлись к полюсам веретена, вокруг каждой группы дочерних хромосом образуется новая ядерная оболочка, которая, возможно, строится из материала одной из мембранных систем цитоплазмы. На этой стадии, носящей название Телофазы, животные клетки обычно делятся путем образования перетяжки, а растительные — путем образования клеточной пластинки. На этом, т. е. на формировании двух дочерних клеток, процесс клеточного деления заканчивается.
Генетическая информация клетки содержится в основном в Хромосомах ядра. Многие детали организации хромосом не выяснены. В так называемой Интерфазе, или метаболической стадии, хромосомный материал обыкновенно мало доступен для наблюдения. Иногда обнаруживаются части хромосом, не подвергшиеся характерным изменениям, сопровождающим митоз. Часто один или несколько таких участков хромосом бывают связаны с Ядрышком, обычно хорошо видимым в метаболически активном ядре.
В растительных и животных клетках между канальцами и пузырьками эндоплазматического ретикулума разбросаны митохондрии. Эти сферические и трубковидные образования также имеют двойную пограничную мембрану, внутренний слой которой образует ряд складок в виде поперечных перегородок, или Крист. В митохондриях протекает большинство реакций, связанных с клеточным дыханием, в том числе образование аденозин - трнфосфата.
Самым заметным образованием внутри большинства клеток является, конечно, ядро. Новейшие электронные микрофотографии показывают, что ядро отделено от цитоплазмы двойной мембраной, которую можно называть Ядерной оболочкой. Внутренняя мембрана, по-видимому, окружает содержимое ядра наподобие мешка. Однако наружная мембрана без перерыва переходит в цитоплазматическую мембранную систему, которая может быть выражена в большей или меньшей степени.
Как растительные, так и животные клетки, по-видимому, имеют наружную оболочку, называемую Плазматической мембраной, которая обладает важным свойством избирательной проницаемости. Как показывают данные физических, химических и биологических исследований, эта мембрана представляет собой сложную структуру, состоящую из белковых и липидных молекул, расположенных слоями.