



Эволюция живых существ может быть понята только в контексте геологического времени.








Эволюция живых существ может быть понята только в контексте геологического времени.

К концу мела, 65 млн. лет назад, абсолютно все группы динозавров вымерли. Вместе с ними исчезли мозазавры, плезиозавры, птерозавры и целый ряд других наземных и морских животных, в том числе аммониты и белемниты. Вымерло 16% семейств морских животных и 18% семейств наземных позвоночных.

В статье рассказывается о наиболее интересных фактах из жизни древних динозавров. Они были самыми крупными из когда-либо живших на суше существ. В мезозойскую эру эти гигантские ящеры безраздельно властвовали на всей Земле. Затем они полностью вымерли, а по земле распространились наши предки — млекопитающие. Но палеонтологи до сих пор продолжают раскапывать все новые и новые окаменелости этих знаменитых ящеров.

Растения и животные произошли от одного примитивного предка в ходе длительной эволюции. Случилось это несколько миллиардов лет назад.
Биогеография изучает биосферу и географическое распределение организмов и их сообществ (биоценозов) в зависимости от условий обитания и геологических особенностей местности, а также характер фауны и флоры отдельных территорий. Биогеография тесно взаимодействует с биологией и экологией. Составными частями в нее входят зоогеография и фитогеография, предметом изучения которых являются соответственно животные и растения.

Батиметрия занимается изучением рельефа подводных частей морей и океанов. Палеобатиметрия изучает подводные глубины, существовавшие в прошлые геологические эпохи. Условия жизни организмов в морской среде позволяют разделить подводные части океанов в вертикальном направлении на области, различающиеся глубиной и, соответственно, давлением и освещенностью.
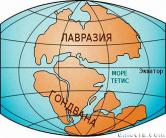
Историческая геология, используя принципы и методы геологии, изучает историю и законы развития Земли со времен образования земной коры в гадейскую эру до нашего времени. Задачи исторической геологии — восстановление хода эволюции земной поверхности и населяющего ее органического мира, а также выяснение истории изменений структуры земной коры.

Бактерии постепенно осваивают кислородный фотосинтез, хотя существенные результаты это даст только в следующую, протерозойскую эру. В конце неоархея находки следов жизни становятся все более распространенными. Среди них стоит отметить группу Малмани, плато Гаап, формации Науга, Когелбеен, Гамохаан и другие.

В неоархее бактериальная жизнь продолжает господствовать в древних земных океанах. Наиболее известные местонахождения следов жизни — это формация Джирина, окрестности огромного стратовулкана Бэк Ривер в Канаде, формация Чешир, субгруппа Шмидтсдриф и другие.

Несмотря на то, что в неоархее жизнь еще не выходила за пределы бактериальных форм, количество находок микрофоссилий и строматолитов постоянно увеличивается. Это такие известные местонахождения, как формация Тумбиана, Маддина, Манджери и находки кратона Дхарвар в Южной Индии.
Цитологический механизм кроссинговера с точностью не известен. Предполагается, что в профазе первого деления мейоза во время синапсиса, когда хромосомы обвивают друг друга, хроматиды разрываются и затем концы их фрагментов вновь соединяются. Если соединятся несестринские хроматиды, произойдет кроссинговер. Кроссинговер происходит с различной частотой в разных участках хромосомы. Вблизи от центромеры и у самых концов хромосом кроссинговера не бывает или он наблюдается редко.
Скрещивая формы, различающиеся по нескольким признакам и гомозиготные по факторам, определяющим эти признаки, Мендель нашел, что факторы различных исследованных им признаков передаются независимо друг от друга.

Единицы наследственности, постулированные Менделем и впоследствии названные Генами, были идентифицированы как специфические участки хромосомы; это отрезки между двумя ближайшими точками кроссинговера К Более поздние исследования, особенно в области биохимической генетики микроорганизмов, привели к другим определениям гена. Например, общепризнано, что специфические гены контролируют образование специфических ферментов.

В процессе репликации тем не менее происходят ошибки; в результате создается изменчивость, обеспечивающая материал для действия отбора. Вообще говоря, анализ природы изменчивости и передачи измененных признаков из поколения в поколение служит единственным средством изучения механизма наследственности. При исследовании характера изменчивости у живых организмов оказывается, что одни организмы, по-видимому, более изменчивы, чем другие.
Во втором делении мейоза хромосомы ведут себя так же, как в митозе, с той лишь разницей, что в результате происшедшего кроссинговера Хроматиды, Прикрепленные к центромере, оказываются не идентичными. Второе деление, при котором происходит деление центромер, ведет к образованию из одного ауксоцита четырех клеток.
По-видимому, на стадии удвоения хромосом тонкие хроматиды в бивалентах разрываются и вновь соединяются концами в местах разрывов. Будучи тесно сближены и перекручены, они часто воссоединяются в несестринских комбинациях, т. е. вместо воссоединения частей сестринских хроматид после разрыва могут соединяться части материнской и отцовской хроматид. Это явление известно под названием Кроссинговера.