



Эволюция живых существ может быть понята только в контексте геологического времени.








Эволюция живых существ может быть понята только в контексте геологического времени.

К концу мела, 65 млн. лет назад, абсолютно все группы динозавров вымерли. Вместе с ними исчезли мозазавры, плезиозавры, птерозавры и целый ряд других наземных и морских животных, в том числе аммониты и белемниты. Вымерло 16% семейств морских животных и 18% семейств наземных позвоночных.

В статье рассказывается о наиболее интересных фактах из жизни древних динозавров. Они были самыми крупными из когда-либо живших на суше существ. В мезозойскую эру эти гигантские ящеры безраздельно властвовали на всей Земле. Затем они полностью вымерли, а по земле распространились наши предки — млекопитающие. Но палеонтологи до сих пор продолжают раскапывать все новые и новые окаменелости этих знаменитых ящеров.

Растения и животные произошли от одного примитивного предка в ходе длительной эволюции. Случилось это несколько миллиардов лет назад.
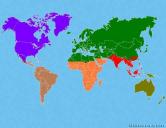
Биогеография изучает биосферу и географическое распределение организмов и их сообществ (биоценозов) в зависимости от условий обитания и геологических особенностей местности, а также характер фауны и флоры отдельных территорий. Биогеография тесно взаимодействует с биологией и экологией. Составными частями в нее входят зоогеография и фитогеография, предметом изучения которых являются соответственно животные и растения.

Батиметрия занимается изучением рельефа подводных частей морей и океанов. Палеобатиметрия изучает подводные глубины, существовавшие в прошлые геологические эпохи. Условия жизни организмов в морской среде позволяют разделить подводные части океанов в вертикальном направлении на области, различающиеся глубиной и, соответственно, давлением и освещенностью.
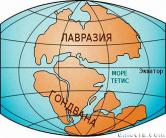
Историческая геология, используя принципы и методы геологии, изучает историю и законы развития Земли со времен образования земной коры в гадейскую эру до нашего времени. Задачи исторической геологии — восстановление хода эволюции земной поверхности и населяющего ее органического мира, а также выяснение истории изменений структуры земной коры.

Бактерии постепенно осваивают кислородный фотосинтез, хотя существенные результаты это даст только в следующую, протерозойскую эру. В конце неоархея находки следов жизни становятся все более распространенными. Среди них стоит отметить группу Малмани, плато Гаап, формации Науга, Когелбеен, Гамохаан и другие.

В неоархее бактериальная жизнь продолжает господствовать в древних земных океанах. Наиболее известные местонахождения следов жизни — это формация Джирина, окрестности огромного стратовулкана Бэк Ривер в Канаде, формация Чешир, субгруппа Шмидтсдриф и другие.

Несмотря на то, что в неоархее жизнь еще не выходила за пределы бактериальных форм, количество находок микрофоссилий и строматолитов постоянно увеличивается. Это такие известные местонахождения, как формация Тумбиана, Маддина, Манджери и находки кратона Дхарвар в Южной Индии.
У раздельнополых животных и растений наблюдается особый тип сцепления. У этих организмов, имеющих специальные Поло- Вые хромосомы, отличные от остальных хромосом, один пол обычно имеет две гомологичные половые хромосомы. У другого пола имеется лишь одна хромосома, гомологичная этим двум, вторая же либо отсутствует, либо отличается от первой и лишь отчасти гомологична ей. У дрозофилы и человека женский пол является Гомогаметным , а мужской пол — Гетерогаметным .
Открытие методов искусственного получения мутаций у легко выращиваемых организмов с относительно коротким жизненным циклом позволило тщательно изучить явление сцепления и группы сцепления, т. е. хромосомы. Если принять, что частота кроссинговера между двумя факторами пропорциональна расстоянию между ними, то можно определять расстояния и порядок расположения генов вдоль хромосомы. Можно построить для хромосомы карту сцепления, основанную на данных о рекомбинациях.
Каким же образом возникает рецессивность? На этот счет существует несколько гипотез, из которых в настоящее время трудно сделать выбор, хотя все они, вероятно, содержат элементы истины. Фишер высказал предположение, что мутации не обязательно должны быть полностью рецессивными при своем первом появлении. Поэтому они, как правило, будут неблагоприятны для организма, если только другие гены-модификаторы, расположенные в иных локусах, не уменьшат вредных последствий их проявления.
В контролируемых скрещиваниях часто удается идентифицировать специфические гены, ответственные за определенные признаки, проявляющие дискретную изменчивость. Наиболее очевидная особенность генов с таким четким действием состоит в том, что они изменяются, или Мутируют; в самом деле, ведь только это и позволяет обнаружить их существование.
В то же время во многих случаях проявление одного гена маскируется действием другого, находящегося в другом локусе. Это явление носит название Эпистаза; эпистатический ген маскирует или предотвращает выражение Гипостатического гена. Например, ген белой окраски у леггорнов является элистатическим по отношению к многим генам, определяющим любую другую окраску и ее распределение. При наличии соответствующего фактора никакая окраска, кроме белой, проявиться не может.
Кроосинговер подвержен влиянию факторов среды, например температуры, и находится также под генетическим контролем. Механизм генетического контроля довольно сложен и еще не вполне ясен. По-видимому, гены определяют возможную длительность синапсиса и локализацию перекрестов; существуют гены, действие которых состоит в полном исключении синапсиса. Следовало бы ожидать, что и другие генетические аномалии, возникающие в процессе мейоза, повлияют на кроссинговер.