



Эволюция живых существ может быть понята только в контексте геологического времени.








Эволюция живых существ может быть понята только в контексте геологического времени.

К концу мела, 65 млн. лет назад, абсолютно все группы динозавров вымерли. Вместе с ними исчезли мозазавры, плезиозавры, птерозавры и целый ряд других наземных и морских животных, в том числе аммониты и белемниты. Вымерло 16% семейств морских животных и 18% семейств наземных позвоночных.

В статье рассказывается о наиболее интересных фактах из жизни древних динозавров. Они были самыми крупными из когда-либо живших на суше существ. В мезозойскую эру эти гигантские ящеры безраздельно властвовали на всей Земле. Затем они полностью вымерли, а по земле распространились наши предки — млекопитающие. Но палеонтологи до сих пор продолжают раскапывать все новые и новые окаменелости этих знаменитых ящеров.

Растения и животные произошли от одного примитивного предка в ходе длительной эволюции. Случилось это несколько миллиардов лет назад.
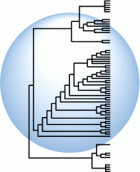
В основе создания всех систематик и классификаций лежит сравнительный анализ связанных родством видов и оценка их сходства и различия по конкретным признакам. И классификация и филогения строятся на изучении естественных групп, сложившихся в природе, имеющих наборы признаков, которые можно обнаружить у их общих предков.

“Рогатое лицо”. Филогенетическая таксономия определяет цератопсов как всех маргиноцефалов, более близко связанных с трицератопсом, чем с пахицефалозавром. Это рогатые растительноядные динозавры, существовавшие в юре и мелу.

“Рыбо-ящеры”. Существовали со раннего триаса до начала позднего мела, расцвета достигли в позднем триасе и ранней юре, затем начали угасать. Со средней юры и до позднего мела существовало уже только несколько семейств. Практически вымерли задолго до конца мела, до начала позднего мела дожил только один род - Platypterygius.
“Ужасные ящеры”. Впервые название использовал Оуэн в своей пересмотренной в 1842 г. для печати версии лекции 1841 г. Кладистический анализ подтверждает динозавров как единую и целостную группу. Она определяется как самый поздний общий предок современных птиц (ящеротазовые) и трицератопса (птицетазовый ящер) и все его потомки. Иногда вместо птиц и трицератопса используются мегалозавр и игуанодон, но это все равно определяет одну и ту же группу.

Возникли, возможно, на границе среднего и позднего девона, 380 млн. лет назад. Окаменелости четвероногообразных известны из позднего девона Гренландии, России, Австралии. Обнаруженные следы тетрапода также известны из раннего–среднего девона Австралии и среднего–позднего девона Бразилии, правда, пальцы на этих отпечатках не видны.

Кембрий. Аномалокариды появились в начале кембрия. Найдены в Австралии, Канаде, Китае, Гренландии и Северной Америке. Это первые всемирно распространенные крупные, активно плавающие хищники, охотящиеся используя зрение, с колючими хватательными придатками и сверхъестественными круглыми ртами, усаженными зубами.
Кроме этих механизмов сохранения или увеличения существующего числа хромосом, у организмов, очевидно, должны существовать механизмы для его уменьшения. Механизмы уменьшения большого числа хромосом, наблюдающегося при эндополиплоидии, мало изучены, но есть данные о существовании таких механизмов у насекомых и у растений. У огромного большинства организмов имеется общий механизм уменьшения зиготического числа хромосом до гаме - тического; этот механизм называется Мейозом .

После того как хромосомы разошлись к полюсам веретена, вокруг каждой группы дочерних хромосом образуется новая ядерная оболочка, которая, возможно, строится из материала одной из мембранных систем цитоплазмы. На этой стадии, носящей название Телофазы, животные клетки обычно делятся путем образования перетяжки, а растительные — путем образования клеточной пластинки. На этом, т. е. на формировании двух дочерних клеток, процесс клеточного деления заканчивается.
Генетическая информация клетки содержится в основном в Хромосомах ядра. Многие детали организации хромосом не выяснены. В так называемой Интерфазе, или метаболической стадии, хромосомный материал обыкновенно мало доступен для наблюдения. Иногда обнаруживаются части хромосом, не подвергшиеся характерным изменениям, сопровождающим митоз. Часто один или несколько таких участков хромосом бывают связаны с Ядрышком, обычно хорошо видимым в метаболически активном ядре.
В растительных и животных клетках между канальцами и пузырьками эндоплазматического ретикулума разбросаны митохондрии. Эти сферические и трубковидные образования также имеют двойную пограничную мембрану, внутренний слой которой образует ряд складок в виде поперечных перегородок, или Крист. В митохондриях протекает большинство реакций, связанных с клеточным дыханием, в том числе образование аденозин - трнфосфата.
Самым заметным образованием внутри большинства клеток является, конечно, ядро. Новейшие электронные микрофотографии показывают, что ядро отделено от цитоплазмы двойной мембраной, которую можно называть Ядерной оболочкой. Внутренняя мембрана, по-видимому, окружает содержимое ядра наподобие мешка. Однако наружная мембрана без перерыва переходит в цитоплазматическую мембранную систему, которая может быть выражена в большей или меньшей степени.
Как растительные, так и животные клетки, по-видимому, имеют наружную оболочку, называемую Плазматической мембраной, которая обладает важным свойством избирательной проницаемости. Как показывают данные физических, химических и биологических исследований, эта мембрана представляет собой сложную структуру, состоящую из белковых и липидных молекул, расположенных слоями.