



Эволюция живых существ может быть понята только в контексте геологического времени.








Эволюция живых существ может быть понята только в контексте геологического времени.

К концу мела, 65 млн. лет назад, абсолютно все группы динозавров вымерли. Вместе с ними исчезли мозазавры, плезиозавры, птерозавры и целый ряд других наземных и морских животных, в том числе аммониты и белемниты. Вымерло 16% семейств морских животных и 18% семейств наземных позвоночных.

В статье рассказывается о наиболее интересных фактах из жизни древних динозавров. Они были самыми крупными из когда-либо живших на суше существ. В мезозойскую эру эти гигантские ящеры безраздельно властвовали на всей Земле. Затем они полностью вымерли, а по земле распространились наши предки — млекопитающие. Но палеонтологи до сих пор продолжают раскапывать все новые и новые окаменелости этих знаменитых ящеров.

Растения и животные произошли от одного примитивного предка в ходе длительной эволюции. Случилось это несколько миллиардов лет назад.
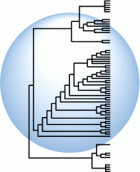
В основе создания всех систематик и классификаций лежит сравнительный анализ связанных родством видов и оценка их сходства и различия по конкретным признакам. И классификация и филогения строятся на изучении естественных групп, сложившихся в природе, имеющих наборы признаков, которые можно обнаружить у их общих предков.

“Рогатое лицо”. Филогенетическая таксономия определяет цератопсов как всех маргиноцефалов, более близко связанных с трицератопсом, чем с пахицефалозавром. Это рогатые растительноядные динозавры, существовавшие в юре и мелу.

“Рыбо-ящеры”. Существовали со раннего триаса до начала позднего мела, расцвета достигли в позднем триасе и ранней юре, затем начали угасать. Со средней юры и до позднего мела существовало уже только несколько семейств. Практически вымерли задолго до конца мела, до начала позднего мела дожил только один род - Platypterygius.
“Ужасные ящеры”. Впервые название использовал Оуэн в своей пересмотренной в 1842 г. для печати версии лекции 1841 г. Кладистический анализ подтверждает динозавров как единую и целостную группу. Она определяется как самый поздний общий предок современных птиц (ящеротазовые) и трицератопса (птицетазовый ящер) и все его потомки. Иногда вместо птиц и трицератопса используются мегалозавр и игуанодон, но это все равно определяет одну и ту же группу.

Возникли, возможно, на границе среднего и позднего девона, 380 млн. лет назад. Окаменелости четвероногообразных известны из позднего девона Гренландии, России, Австралии. Обнаруженные следы тетрапода также известны из раннего–среднего девона Австралии и среднего–позднего девона Бразилии, правда, пальцы на этих отпечатках не видны.

Кембрий. Аномалокариды появились в начале кембрия. Найдены в Австралии, Канаде, Китае, Гренландии и Северной Америке. Это первые всемирно распространенные крупные, активно плавающие хищники, охотящиеся используя зрение, с колючими хватательными придатками и сверхъестественными круглыми ртами, усаженными зубами.
Цитологический механизм кроссинговера с точностью не известен. Предполагается, что в профазе первого деления мейоза во время синапсиса, когда хромосомы обвивают друг друга, хроматиды разрываются и затем концы их фрагментов вновь соединяются. Если соединятся несестринские хроматиды, произойдет кроссинговер. Кроссинговер происходит с различной частотой в разных участках хромосомы. Вблизи от центромеры и у самых концов хромосом кроссинговера не бывает или он наблюдается редко.
Скрещивая формы, различающиеся по нескольким признакам и гомозиготные по факторам, определяющим эти признаки, Мендель нашел, что факторы различных исследованных им признаков передаются независимо друг от друга.

Единицы наследственности, постулированные Менделем и впоследствии названные Генами, были идентифицированы как специфические участки хромосомы; это отрезки между двумя ближайшими точками кроссинговера К Более поздние исследования, особенно в области биохимической генетики микроорганизмов, привели к другим определениям гена. Например, общепризнано, что специфические гены контролируют образование специфических ферментов.

В процессе репликации тем не менее происходят ошибки; в результате создается изменчивость, обеспечивающая материал для действия отбора. Вообще говоря, анализ природы изменчивости и передачи измененных признаков из поколения в поколение служит единственным средством изучения механизма наследственности. При исследовании характера изменчивости у живых организмов оказывается, что одни организмы, по-видимому, более изменчивы, чем другие.
Во втором делении мейоза хромосомы ведут себя так же, как в митозе, с той лишь разницей, что в результате происшедшего кроссинговера Хроматиды, Прикрепленные к центромере, оказываются не идентичными. Второе деление, при котором происходит деление центромер, ведет к образованию из одного ауксоцита четырех клеток.
По-видимому, на стадии удвоения хромосом тонкие хроматиды в бивалентах разрываются и вновь соединяются концами в местах разрывов. Будучи тесно сближены и перекручены, они часто воссоединяются в несестринских комбинациях, т. е. вместо воссоединения частей сестринских хроматид после разрыва могут соединяться части материнской и отцовской хроматид. Это явление известно под названием Кроссинговера.