



Эволюция живых существ может быть понята только в контексте геологического времени.








Эволюция живых существ может быть понята только в контексте геологического времени.

К концу мела, 65 млн. лет назад, абсолютно все группы динозавров вымерли. Вместе с ними исчезли мозазавры, плезиозавры, птерозавры и целый ряд других наземных и морских животных, в том числе аммониты и белемниты. Вымерло 16% семейств морских животных и 18% семейств наземных позвоночных.

В статье рассказывается о наиболее интересных фактах из жизни древних динозавров. Они были самыми крупными из когда-либо живших на суше существ. В мезозойскую эру эти гигантские ящеры безраздельно властвовали на всей Земле. Затем они полностью вымерли, а по земле распространились наши предки — млекопитающие. Но палеонтологи до сих пор продолжают раскапывать все новые и новые окаменелости этих знаменитых ящеров.

Растения и животные произошли от одного примитивного предка в ходе длительной эволюции. Случилось это несколько миллиардов лет назад.
Биогеография изучает биосферу и географическое распределение организмов и их сообществ (биоценозов) в зависимости от условий обитания и геологических особенностей местности, а также характер фауны и флоры отдельных территорий. Биогеография тесно взаимодействует с биологией и экологией. Составными частями в нее входят зоогеография и фитогеография, предметом изучения которых являются соответственно животные и растения.

Батиметрия занимается изучением рельефа подводных частей морей и океанов. Палеобатиметрия изучает подводные глубины, существовавшие в прошлые геологические эпохи. Условия жизни организмов в морской среде позволяют разделить подводные части океанов в вертикальном направлении на области, различающиеся глубиной и, соответственно, давлением и освещенностью.
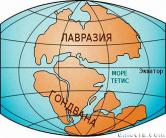
Историческая геология, используя принципы и методы геологии, изучает историю и законы развития Земли со времен образования земной коры в гадейскую эру до нашего времени. Задачи исторической геологии — восстановление хода эволюции земной поверхности и населяющего ее органического мира, а также выяснение истории изменений структуры земной коры.

Бактерии постепенно осваивают кислородный фотосинтез, хотя существенные результаты это даст только в следующую, протерозойскую эру. В конце неоархея находки следов жизни становятся все более распространенными. Среди них стоит отметить группу Малмани, плато Гаап, формации Науга, Когелбеен, Гамохаан и другие.

В неоархее бактериальная жизнь продолжает господствовать в древних земных океанах. Наиболее известные местонахождения следов жизни — это формация Джирина, окрестности огромного стратовулкана Бэк Ривер в Канаде, формация Чешир, субгруппа Шмидтсдриф и другие.

Несмотря на то, что в неоархее жизнь еще не выходила за пределы бактериальных форм, количество находок микрофоссилий и строматолитов постоянно увеличивается. Это такие известные местонахождения, как формация Тумбиана, Маддина, Манджери и находки кратона Дхарвар в Южной Индии.
Митоз был охарактеризован как механизм, обеспечивающий равное распределение генетической информации между дочерними клетками в ходе клеточного деления. То, что митоз может достигать этого результата, легко доказывается целым рядом способов. Если, например, зиготическому ядру стрекозы Platyc- Nemis дать возможность пройти через 7 делений, а затем все дочерние клетки, кроме одной, убить с помощью узкого пучка ультрафиолетовых лучей, то из этой клетки все-таки разовьется нормальный эмбрион.
Простой цикл «рост — фрагментация — рост», постулированный в качестве наиболее примитивной формы развития, претерпел в результате отбора разнообразные изменения. Примером сложного цикла развития может служить жизненный цикл простейшего Plasmodium — возбудителя малярии. Спорозоиты Plasmodium в организме комара мигрируют в слюнные железы, а затем, попадая при укусе в кровяное русло человека, способны проникать в клетки определенного типа.
Исследования последних лет внесли уточнения и поправки относительно принципов деления исходной группы. Уход одной или нескольких матрилиний—лишь одна, притом наиболее простая модель. Чаще всего дело обстоит не так просто. Наблюдения за группой резусов, живущих на о-ве Кайо-Сантьяго, показали, что особи четырех, из шести генеалогий распределились между двумя дочерними группами. Каждая матрилиния разделилась приблизительно на равные части, точно по линиям доминирования.
Матери обеспечивают высокие ранги своим детенышам не путем воспитания в них высокоагрессивных особей, а благодаря формированию устойчивого стереотипа подчиненности у детенышей низкоранговых самок. Установление иерархических отношений в группе указанными путями является важным фактором, способствующим сохранению групповой стабильности. Реверсия рангов старшей и младшей сестер во многом связана с поведением матери, чаще принимающей сторону младшей в конфликтных ситуациях.
Тонкотелые обезьяны. Роды у лангуров в естественных условиях наблюдал Оппенгеймер. Он описывает, как за роженицей внимательно следили две взрослые самки — вначале издалека, сидя на дереве, затем спустились на землю и сели рядом с рожающей особью. Представители другого вида лангуров — Р. nemaeus, напротив, совершенно индифферентно отнеслись к процессу родов: не заинтересовал их сразу и новорожденный детеныш. Лишь неделю спустя самка-мать и детеныш оказались в фокусе внимания членов группы.
Широконосые обезьяны. У цебид ревунов роды происходят в присутствии самца и старших детенышей. У Callicebus moloch с первых мгновений появления детенышей на свет самец берет их на руки, носит и проявляет все больше заботы о них со временем. Аналогичное поведение самца-отца отмечено у мармазеток и тамаринов. У видов широконосых обезьян, живущих новорожденного детеныша. К сожалению, данные, описывающие появление детеныша в естественных условиях, пока фрагментарны.