



Эволюция живых существ может быть понята только в контексте геологического времени.








Эволюция живых существ может быть понята только в контексте геологического времени.

К концу мела, 65 млн. лет назад, абсолютно все группы динозавров вымерли. Вместе с ними исчезли мозазавры, плезиозавры, птерозавры и целый ряд других наземных и морских животных, в том числе аммониты и белемниты. Вымерло 16% семейств морских животных и 18% семейств наземных позвоночных.

В статье рассказывается о наиболее интересных фактах из жизни древних динозавров. Они были самыми крупными из когда-либо живших на суше существ. В мезозойскую эру эти гигантские ящеры безраздельно властвовали на всей Земле. Затем они полностью вымерли, а по земле распространились наши предки — млекопитающие. Но палеонтологи до сих пор продолжают раскапывать все новые и новые окаменелости этих знаменитых ящеров.

Растения и животные произошли от одного примитивного предка в ходе длительной эволюции. Случилось это несколько миллиардов лет назад.
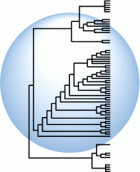
В основе создания всех систематик и классификаций лежит сравнительный анализ связанных родством видов и оценка их сходства и различия по конкретным признакам. И классификация и филогения строятся на изучении естественных групп, сложившихся в природе, имеющих наборы признаков, которые можно обнаружить у их общих предков.

“Рогатое лицо”. Филогенетическая таксономия определяет цератопсов как всех маргиноцефалов, более близко связанных с трицератопсом, чем с пахицефалозавром. Это рогатые растительноядные динозавры, существовавшие в юре и мелу.

“Рыбо-ящеры”. Существовали со раннего триаса до начала позднего мела, расцвета достигли в позднем триасе и ранней юре, затем начали угасать. Со средней юры и до позднего мела существовало уже только несколько семейств. Практически вымерли задолго до конца мела, до начала позднего мела дожил только один род - Platypterygius.
“Ужасные ящеры”. Впервые название использовал Оуэн в своей пересмотренной в 1842 г. для печати версии лекции 1841 г. Кладистический анализ подтверждает динозавров как единую и целостную группу. Она определяется как самый поздний общий предок современных птиц (ящеротазовые) и трицератопса (птицетазовый ящер) и все его потомки. Иногда вместо птиц и трицератопса используются мегалозавр и игуанодон, но это все равно определяет одну и ту же группу.

Возникли, возможно, на границе среднего и позднего девона, 380 млн. лет назад. Окаменелости четвероногообразных известны из позднего девона Гренландии, России, Австралии. Обнаруженные следы тетрапода также известны из раннего–среднего девона Австралии и среднего–позднего девона Бразилии, правда, пальцы на этих отпечатках не видны.

Кембрий. Аномалокариды появились в начале кембрия. Найдены в Австралии, Канаде, Китае, Гренландии и Северной Америке. Это первые всемирно распространенные крупные, активно плавающие хищники, охотящиеся используя зрение, с колючими хватательными придатками и сверхъестественными круглыми ртами, усаженными зубами.
Определение признака многими генами обусловливает такую стабильность фенотипического выражения, какой может и не быть, если признак определяется лишь одним геном. Например, единичная мутация вряд ли серьезно нарушит проявление признака, зависящего, скажем, от аддитивных эффектов 35 локусов. Напротив, единичная мутация г->/ в гене, подавляющем окраску у лука, приведет к образованию белой луковицы, а не красной или желтой.
Обычно бывает трудно разделить генетические и средовые компоненты изменчивости и исследовать отдельно ту ее часть, которая обусловлена генетически. Для изучения и разделения этих компонентов были разработаны различные экспериментальные методы и использован довольно сложный математический аппарат. Например, один из способов оценки роли этих двух компонентов основан на уменьшении генетического компонента до пренебрежимо малой величины.
Переходя к изучению наследования признаков, подверженных непрерывной изменчивости, мы сталкиваемся с еще более трудными проблемами. Обычно невозможно бывает выявить специфические гены, контролирующие, определенные признаки. Как уже упоминалось, существует, по-видимому, непрерывный спектр изменчивости от типично качественной до чисто количественной.
Генетически все это выглядит так, как если бы кроссинговер был подавлен. Однако на самом деле происходит только элиминация его продуктов. Последствия зависят от числа перекрестов в инверсионной петле и от их распределения между хроматидами бивалента. Так как у гетерозигот гены, лежащие в инверсионной петле, при известных условиях оказываются эффективно сцепленными, инверсии будут подробнее рассмотрены в связи с рекомбинацией.

Фразеологизм "нем как рыба" давно перестал использоваться в научных кругах, так как биологи установили, что рыбы ещё как "разговаривают" между собой, а некоторые из них даже повышают голос. Но учёные из университета Майами открыли миру новую удивительную тайну: оказывается, не только рыбы, но и их личинки издают звуки.
Делеции и дупликации. Роль в эволюции утраты или добавления хромосомного материала еще очень неясна. В гомозиготном состоянии делеции обычно летальны. Делеции служат полезным инструментом при построении хромосомных карт; их эволюционное значение в настоящее время неизвестно. Значение дупликаций состоит в том, что они составляют возможный цитогенетический механизм увеличения общего количества генетического материала. Как изменялось это количество в ходе эволюции — мы не знаем.