



Эволюция живых существ может быть понята только в контексте геологического времени.








Эволюция живых существ может быть понята только в контексте геологического времени.

К концу мела, 65 млн. лет назад, абсолютно все группы динозавров вымерли. Вместе с ними исчезли мозазавры, плезиозавры, птерозавры и целый ряд других наземных и морских животных, в том числе аммониты и белемниты. Вымерло 16% семейств морских животных и 18% семейств наземных позвоночных.

В статье рассказывается о наиболее интересных фактах из жизни древних динозавров. Они были самыми крупными из когда-либо живших на суше существ. В мезозойскую эру эти гигантские ящеры безраздельно властвовали на всей Земле. Затем они полностью вымерли, а по земле распространились наши предки — млекопитающие. Но палеонтологи до сих пор продолжают раскапывать все новые и новые окаменелости этих знаменитых ящеров.

Растения и животные произошли от одного примитивного предка в ходе длительной эволюции. Случилось это несколько миллиардов лет назад.
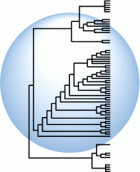
В основе создания всех систематик и классификаций лежит сравнительный анализ связанных родством видов и оценка их сходства и различия по конкретным признакам. И классификация и филогения строятся на изучении естественных групп, сложившихся в природе, имеющих наборы признаков, которые можно обнаружить у их общих предков.

“Рогатое лицо”. Филогенетическая таксономия определяет цератопсов как всех маргиноцефалов, более близко связанных с трицератопсом, чем с пахицефалозавром. Это рогатые растительноядные динозавры, существовавшие в юре и мелу.

“Рыбо-ящеры”. Существовали со раннего триаса до начала позднего мела, расцвета достигли в позднем триасе и ранней юре, затем начали угасать. Со средней юры и до позднего мела существовало уже только несколько семейств. Практически вымерли задолго до конца мела, до начала позднего мела дожил только один род - Platypterygius.
“Ужасные ящеры”. Впервые название использовал Оуэн в своей пересмотренной в 1842 г. для печати версии лекции 1841 г. Кладистический анализ подтверждает динозавров как единую и целостную группу. Она определяется как самый поздний общий предок современных птиц (ящеротазовые) и трицератопса (птицетазовый ящер) и все его потомки. Иногда вместо птиц и трицератопса используются мегалозавр и игуанодон, но это все равно определяет одну и ту же группу.

Возникли, возможно, на границе среднего и позднего девона, 380 млн. лет назад. Окаменелости четвероногообразных известны из позднего девона Гренландии, России, Австралии. Обнаруженные следы тетрапода также известны из раннего–среднего девона Австралии и среднего–позднего девона Бразилии, правда, пальцы на этих отпечатках не видны.

Кембрий. Аномалокариды появились в начале кембрия. Найдены в Австралии, Канаде, Китае, Гренландии и Северной Америке. Это первые всемирно распространенные крупные, активно плавающие хищники, охотящиеся используя зрение, с колючими хватательными придатками и сверхъестественными круглыми ртами, усаженными зубами.
Митоз был охарактеризован как механизм, обеспечивающий равное распределение генетической информации между дочерними клетками в ходе клеточного деления. То, что митоз может достигать этого результата, легко доказывается целым рядом способов. Если, например, зиготическому ядру стрекозы Platyc- Nemis дать возможность пройти через 7 делений, а затем все дочерние клетки, кроме одной, убить с помощью узкого пучка ультрафиолетовых лучей, то из этой клетки все-таки разовьется нормальный эмбрион.
Простой цикл «рост — фрагментация — рост», постулированный в качестве наиболее примитивной формы развития, претерпел в результате отбора разнообразные изменения. Примером сложного цикла развития может служить жизненный цикл простейшего Plasmodium — возбудителя малярии. Спорозоиты Plasmodium в организме комара мигрируют в слюнные железы, а затем, попадая при укусе в кровяное русло человека, способны проникать в клетки определенного типа.
Исследования последних лет внесли уточнения и поправки относительно принципов деления исходной группы. Уход одной или нескольких матрилиний—лишь одна, притом наиболее простая модель. Чаще всего дело обстоит не так просто. Наблюдения за группой резусов, живущих на о-ве Кайо-Сантьяго, показали, что особи четырех, из шести генеалогий распределились между двумя дочерними группами. Каждая матрилиния разделилась приблизительно на равные части, точно по линиям доминирования.
Матери обеспечивают высокие ранги своим детенышам не путем воспитания в них высокоагрессивных особей, а благодаря формированию устойчивого стереотипа подчиненности у детенышей низкоранговых самок. Установление иерархических отношений в группе указанными путями является важным фактором, способствующим сохранению групповой стабильности. Реверсия рангов старшей и младшей сестер во многом связана с поведением матери, чаще принимающей сторону младшей в конфликтных ситуациях.
Тонкотелые обезьяны. Роды у лангуров в естественных условиях наблюдал Оппенгеймер. Он описывает, как за роженицей внимательно следили две взрослые самки — вначале издалека, сидя на дереве, затем спустились на землю и сели рядом с рожающей особью. Представители другого вида лангуров — Р. nemaeus, напротив, совершенно индифферентно отнеслись к процессу родов: не заинтересовал их сразу и новорожденный детеныш. Лишь неделю спустя самка-мать и детеныш оказались в фокусе внимания членов группы.
Широконосые обезьяны. У цебид ревунов роды происходят в присутствии самца и старших детенышей. У Callicebus moloch с первых мгновений появления детенышей на свет самец берет их на руки, носит и проявляет все больше заботы о них со временем. Аналогичное поведение самца-отца отмечено у мармазеток и тамаринов. У видов широконосых обезьян, живущих новорожденного детеныша. К сожалению, данные, описывающие появление детеныша в естественных условиях, пока фрагментарны.