



Эволюция живых существ может быть понята только в контексте геологического времени.








Эволюция живых существ может быть понята только в контексте геологического времени.

К концу мела, 65 млн. лет назад, абсолютно все группы динозавров вымерли. Вместе с ними исчезли мозазавры, плезиозавры, птерозавры и целый ряд других наземных и морских животных, в том числе аммониты и белемниты. Вымерло 16% семейств морских животных и 18% семейств наземных позвоночных.

В статье рассказывается о наиболее интересных фактах из жизни древних динозавров. Они были самыми крупными из когда-либо живших на суше существ. В мезозойскую эру эти гигантские ящеры безраздельно властвовали на всей Земле. Затем они полностью вымерли, а по земле распространились наши предки — млекопитающие. Но палеонтологи до сих пор продолжают раскапывать все новые и новые окаменелости этих знаменитых ящеров.

Растения и животные произошли от одного примитивного предка в ходе длительной эволюции. Случилось это несколько миллиардов лет назад.

+ Pleurocaulis rewanense. Плеврокаулис реванский. «Ребристый стебель. Видовое имя от названия стратиграфического слоя Rewan Group». Ранний триас, Австралия (Квинсленд). Известны вегетативные структуры Pleurocaulis rewanense. Протостелический стебель шириной до 20 мм, несущий спирально расположенные круглые или овальные листовые рубцы или продольные борозды 10 мм длиной и 1 мм глубиной, которые могут представлять декортифицированную стадию стебля.

+ Pleuromeia. Плевромейя. Ранний–средний триас, Лавразия, Гондвана (Германия, Франция, Испания, Россия, Каспий, Узбекистан, Австралия, Аргентина, Япония, Китай). Разноспоровая, но стробилы были моноспорангиевыми, содержащими либо микроспоры, либо мегаспоры. Предпочитала околоводные и низинные местности.

Ранний–поздний триас (Scythian–Carnian). Pleuromeiaceae — это семейство вымерших плауновидных растений. Это были небольшие прямые неветвящиеся деревца около одного–двух метров высотой, с прямым стволом без ветвей, со способностью к вторичному росту стебля. Плодоношения у Pleuromeiaceae могут быть и односпоровыми, и двухспоровыми. Спорангии часто погружены в спорофилл.
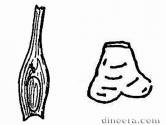
Семейство Takhtajanodoxaceae включает в себя только один род. Имеется лигула с глоссоподием, найденные только у этого семейства. Близки по своему строению к семействам Chaloneriaceae и Isoetaceae.

Поздний карбон (Bashkirian–Kasimovian). Chaloneriaceae — это ранние примитивные члены отряда изоетовых с прямым неветвящимся стеблем и утолщенным, клубнеобразным основанием.

Представители этого порядка являются травянистыми или псевдотравянистыми формами, произошедшими от общих предков с древовидными плауновидными каменноугольного времени в результате приспособления их к климату последующих геологических периодов.
Определение признака многими генами обусловливает такую стабильность фенотипического выражения, какой может и не быть, если признак определяется лишь одним геном. Например, единичная мутация вряд ли серьезно нарушит проявление признака, зависящего, скажем, от аддитивных эффектов 35 локусов. Напротив, единичная мутация г->/ в гене, подавляющем окраску у лука, приведет к образованию белой луковицы, а не красной или желтой.
Обычно бывает трудно разделить генетические и средовые компоненты изменчивости и исследовать отдельно ту ее часть, которая обусловлена генетически. Для изучения и разделения этих компонентов были разработаны различные экспериментальные методы и использован довольно сложный математический аппарат. Например, один из способов оценки роли этих двух компонентов основан на уменьшении генетического компонента до пренебрежимо малой величины.
Переходя к изучению наследования признаков, подверженных непрерывной изменчивости, мы сталкиваемся с еще более трудными проблемами. Обычно невозможно бывает выявить специфические гены, контролирующие, определенные признаки. Как уже упоминалось, существует, по-видимому, непрерывный спектр изменчивости от типично качественной до чисто количественной.
Генетически все это выглядит так, как если бы кроссинговер был подавлен. Однако на самом деле происходит только элиминация его продуктов. Последствия зависят от числа перекрестов в инверсионной петле и от их распределения между хроматидами бивалента. Так как у гетерозигот гены, лежащие в инверсионной петле, при известных условиях оказываются эффективно сцепленными, инверсии будут подробнее рассмотрены в связи с рекомбинацией.

Фразеологизм "нем как рыба" давно перестал использоваться в научных кругах, так как биологи установили, что рыбы ещё как "разговаривают" между собой, а некоторые из них даже повышают голос. Но учёные из университета Майами открыли миру новую удивительную тайну: оказывается, не только рыбы, но и их личинки издают звуки.
Делеции и дупликации. Роль в эволюции утраты или добавления хромосомного материала еще очень неясна. В гомозиготном состоянии делеции обычно летальны. Делеции служат полезным инструментом при построении хромосомных карт; их эволюционное значение в настоящее время неизвестно. Значение дупликаций состоит в том, что они составляют возможный цитогенетический механизм увеличения общего количества генетического материала. Как изменялось это количество в ходе эволюции — мы не знаем.