



Эволюция живых существ может быть понята только в контексте геологического времени.








Эволюция живых существ может быть понята только в контексте геологического времени.

К концу мела, 65 млн. лет назад, абсолютно все группы динозавров вымерли. Вместе с ними исчезли мозазавры, плезиозавры, птерозавры и целый ряд других наземных и морских животных, в том числе аммониты и белемниты. Вымерло 16% семейств морских животных и 18% семейств наземных позвоночных.

В статье рассказывается о наиболее интересных фактах из жизни древних динозавров. Они были самыми крупными из когда-либо живших на суше существ. В мезозойскую эру эти гигантские ящеры безраздельно властвовали на всей Земле. Затем они полностью вымерли, а по земле распространились наши предки — млекопитающие. Но палеонтологи до сих пор продолжают раскапывать все новые и новые окаменелости этих знаменитых ящеров.

Растения и животные произошли от одного примитивного предка в ходе длительной эволюции. Случилось это несколько миллиардов лет назад.

“Разнозубые ящеры”. Поздний триас (Norian)–ранний мел (Barremian), часть Гондваны (Южная Африка, Южная Америка) и все континенты Лавразии (Северный Китай, Европа, запад Северной Америки). Ранние небольшие двуногие травоядные (факультативно всеядные) птицетазовые ящеры. Длина тела обычно меньше 1 м, самый крупный достигал длины 1,2 м, вес тела до 20 кг. Большинство диагностически определенных гетеродонтозаврид происходят из Южной Африки (найдено около 20 экземпляров), где, наряду с Южной Америкой, находился в позднем триасе центр возникновения и распространения всех птицетазовых динозавров.

Сравнение размеров известных гетеродонтозаврид. Большинство таксонов были одинаково малыми, приблизительно 60-80 см длиной и 20-30 см высотой; но два рода были заметно крупнее: Heterodontosaurus tucki был по крайней мере более 100 см длиной и 35 см высотой, и Lycorhinus angustidens был 90-120 см длиной и 25-40 см высотой.
Большой диапазон размеров для Lycorhinus происходит из-за полного отсутствия посткраниального материала. Если у Lycorhinus был крупный череп, как у Abrictosaurus (меньший силуэт) Lycorhinus бы оказался меньше Heterodontosaurus, но если у него малый череп относительно тела, как у Heterodontosaurus, он бы даже был несколько крупнее его. Также, хотя Fruitadens рекламировался в средствах массовой информации, как самый малый известный не-птичий динозавр, он, кажется, был таким же, как пять остальных гетеродонтозавридов
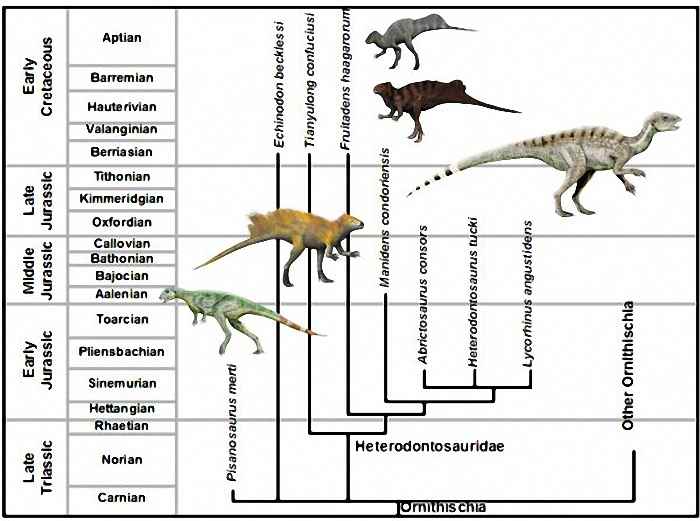
Семейство Heterodontosauridae — это морфологически разнообразная группа динозавров, которая интерпретируется как одно из наиболее базальных семейств ранних орнитисхий, немногочисленное по составу и редкое в ископаемых находках, но довольно широко распространенное почти по всем континентам (за исключением Австралии и Антарктиды). Пангея в юрском периоде представляла собой единое целое, что способствовало их всемирному распространению.
 |
 |
| Палеогеографическая карта распространения гетеродонтозаврид. A, неназванный таксон, формация Laguna Colorada, Аргентина. B, Heterodontosaurus, Abrictosaurus, Lycorhinus, формации Upper Elliot and Clarens, Южная Африка. C, неописанный таксон, формация Kayenta, США. D, Fruitadens, формация Morrison, США. E, Echinodon, формация Purbeck, Англия. F, Tianyulong, формация Yixian, Китай |
Биогеографическое распространение гетеродонтозаврид и филогенетический анализ. Расположение материков соответствует раннему мелу. A - Африка, As - Азия, E - Европа, NA - Северная Америка, SA - Южная Америка. |
Первоначально они были известны из ранней юры Южной Африки, но современные исследования значительно расширили географический и временной диапазоны для этого клэда. Наиболее древней находкой является фрагмент челюсти и изолированные зубы из аргентинской формации Лагуна Колорада (Патагония), датируемые поздним триасом (Norian). Эти остатки имеют развитую морфологию, аналогичную с Heterodontosaurus, включая клыки с зазубринами на переднем и заднем краях, а также щечные, тесно расположенные зубы с высокой кроной и без цингулюма. Наиболее разнообразная фауна гетеродонтозаврид отмечается в ранней юре Южной Африки. Неописанные раннеюрские (Sinemurian–Pliensbachian) гетеродонтозавриды известны также из США, Аризона (формация Кайента), и Мексики; гетеродонтовые зубы были открыты в отложениях поздней юры и раннего мела Испании и Португалии. Позднеюрские и раннемеловые гетеродонтозавриды известны только из Лавразии. Последние представители гетеродонтозаврид вымерли лишь в первой половине мела. Таким образом, эволюционная линия гетеродонтозаврид простиралась более чем на 100 миллионов лет, делая их одной из наиболее долгоживущих групп из всех ранних динозавров.
Состав группы и, соответственно, ее распространение не определены окончательно, в основном из-за фрагментарности некоторых важных находок. Поздние виды из раннего мела имеют достаточно примитивное строение и менее специализированы, их диета включала большее количество беспозвоночных. Среди гетеродонтозаврид только сам Heterodontosaurus известен по полному скелету, большинство других представителей семейства представлены весьма фрагментарно. Поэтому одно из главных свойств семейства — морфология зубов и большинство синапоморфий гетеродонтозаврид описаны по зубам и челюстям, так как это самая ранняя группа динозавров с дифференцированной зубной системой.
Почти все они имели очень большие глаза. Ниже глаз скула выступала по бокам черепа, что свойственно также цератопсам. Скула у гетеродонтозаврид связывает верхнюю челюсть с подвеской, замыкая снизу глазницу и межвисочное отверстие.

Верхняя челюсть Heterodontosaurus tucki
Само название гетеродонтозаврид возникло на основании их сильно различных видов зубов в зубном ряду. Многие имели три вида зубов — острые спереди верхней челюсти, ряды щечных зубов для размельчения растений и между ними пары острых клыков в специальных гнездах. Верхние клыки были более короткими. Характерны долотообразные кроны верхнечелюстных зубов с зазубринами, ограниченными только верхней третью высоты зуба, заметные мезиальные и дистальные гребни на верхнечелюстных зубах. Хотя такие зубы характерны для травоядных, гетеродонтозавры, очевидно, также ели насекомых.
Как и в челюстях всех орнитисхий, передний край предчелюстной кости был беззубым и поддерживал ороговевший клюв (рамфотеку), хотя гетеродонтозавриды имели зубы в задней части предчелюстной кости. У большинства орнитисхий большой пробел, называемый диастемой, отделял эти зубы от зубов на верхнечелюстной кости, но эта диастема была характерно дугообразной у гетеродонтозаврид. Все зубы в нижней челюсти располагались на дентарии, передний край которого также был беззубым (на расстоянии двух альвеол). После клыка в дентарии имелось около 9 зубов.

Heterodontosaurus tucki. Левая верхняя челюсть и правый дентарий (внизу)
На предчелюстной кости имелось 3 зуба. Многие другие базальные орнитисхии имеют 6 или больше зубов, многие заурисхии имеют, по меньшей мере, 4 зуба. У раннеюрских форм первые 2 зуба предчелюстной кости были маленькими и коническими, а третий зуб напоминал клык хищного млекопитающего. Он был стройный, почти прямой с неорнаментированным передним и задним килем. Нижний клык был больше, чем верхний и занимал переднее положение в дентарии. Когда рот был закрыт, он входил в изогнутую диастему на верхней челюсти. У некоторых видов эти клыки были зазубрены на передних и задних краях, у других только на передних. Более поздние формы могли иметь несколько иное расположение клыков. Грани износа показывают, что клыки, в первую очередь, использовались для подрезания пищи, может также для защиты и демонстрации, но слабо подходили для захвата или жевания. В отличие от гетеродонтозаврид, у примитивных амниот и млекопитающих клыки обычно расположены на передней части верхнечелюстной кости.

Heterodontosaurus tucki
Короткие клыки, найденные у всех известных гетеродонтозаврид, сильно напоминают клыки у современных кабарги, пекари и свиней. У многих этих животных клыки являются элементом полового диморфизма и имеются только у самцов, которым могут служить турнирным оружием. Кроме того, возможно, что клыки некоторых гетеродонтозаврид достигали своего нормального размера только у взрослых животных, а не были признаком полового диморфизма.
Как характерные клыки, щечные зубы развитых гетеродонтозаврид были также уникальными среди орнитисхий. Маленькие зубчики находились на краях щечных зубов орнитисхий, чтобы лучше подрезать растительность. Эти зубчики простирались только на верхнюю треть коронки зуба у всех гетеродонтозаврид, а у остальных птицетазовых динозавров они простирались дальше вниз, к корню. Базальные формы (абриктозавр) имели щечные зубы и на верхней челюсти, и на дентарии, структурой они были подобны зубам остальных орнитисхий — широкие промежутки между зубами, низкая крона и сильно развитый поясок, отделяющий крону от корня (цингулюм). У более развитых форм, таких как Lycorhinus и Heterodontosaurus, эти зубы имели форму долота, с намного более высокой кроной и отсутствием цингулюма, поэтому они не имели различия по ширине между кроной и корнем зуба.

Правая верхняя челюсть Heterodontosaurus tucki. А - вид сбоку; В - план коронок, демонстрирующий небольшое перекрывание прилегающих зубов
Эти развитые щечные зубы перекрывались так, чтобы их кроны образовывали непрерывную поверхность, которой можно было бы перемалывать растительную пищу. Зубные ряды были несколько смещены внутрь от края челюстей, оставляя небольшое пространство, которое возможно было ограничено снаружи мышечной щекой, необходимой при таком типе жевания. Край кости выступал вбок, образуя гребни для поддержки мягкой ткани щеки на верхнечелюстной и зубной костях, соответственно выше и ниже зубного ряда. Меловые гадрозавры и цератопсы, в ходе конвергентной эволюции развили схожие зубные батареи. В отличие от гадрозавров, которые имели тысячи постоянно замещаемых зубов, у гетеродонтозаврид замена происходила намного медленней и некоторые экземпляры найдены без замещенных зубов. Но в целом, замена зубов была для них более обычной, чем это предполагалось первоначально. Гипотеза, которую выдвинул Талборн о том, что гетеродонтозавриды на сухой летний период впадали в некое подобие спячки, во время которого и происходила замена зубов, не нашла никакого подтверждения и опровергнута находками замены зубов во всех известных видах гетеродонтозаврид.
Характерно, что у гетеродонтозаврид имелся недостаток малых отверстий на внутренней части костей челюстей, которые, как считается, помогали развитию зубов у других птицетазовых динозавров.
Гетеродонтозавриды также имели уникальное сочленение между дентарием и предентарием, позволяющее нижним челюстям несколько поворачиваться наружу, давая возможность щечным зубам перетирать пищу друг о друга, когда рот был закрыт. Из-за медленной замены зубов, такое измельчение пищи приводило к экстремальному износу зубов, который стирал большинство зубчиков на старых зубах, хотя увеличенная высота кроны давала каждому зубу продолжительный срок службы. Износ зубов указывает на специфический тип прикуса долотовидных верхних и нижних зубов: латеральная поверхность нижних стирается о медиальную поверхность верхних. Эмаль же, напротив, покрывает только латеральную поверхность верхнечелюстных зубов и медиальную поверхность нижнечелюстных, что приводит к образованию устойчивого самозатачивающегося режущего края.

Правая передняя конечность Heterodontosaurus tucki
Гетеродонтозавриды характеризовались необычно длинными передними конечностями, приспособленными для схватывания, а не для поддержки веса тела. Пропорции и анатомия задних конечностей указывают на двуногое передвижение и хорошие способности к бегу. Это подтверждается скелетом Heterodontosaurus tucki, сохранившим шейные позвонки сочлененными друг с другом. Они придают его шее сигмовидную форму кривизны, обычную для двуногих животных. Сочленение затылочного и спинного позвонков свидетельствуют, что тело динозавра при этом располагалось горизонтально.
Хотя длина передних конечностей предполагает хотя бы частичную четвероногость, строение кисти передней конечности ясно приспособлено к схватыванию, а не передвижению и не имело адаптаций для поддержания веса тела. Кроме того, когтевые фаланги передних конечностей были уплощены с боков, как и сами когти, и не годились для ходьбы. У всех четырехногих птицетазовых динозавров (например, игуанодонтид и гадрозаврид) когтевые фаланги наоборот, расширяются в стороны, образуя копытоподобную структуру для лучшего распределения нагрузки.
Передние конечности были весьма длинными для динозавра, достигая 70% от длины задних ног. Хорошо развитый дельтапекторальный гребень плечевой кости (гребень для прикрепления грудных и плечевых мышц) и заметный локтевой отросток (где крепятся разгибающие мышцы предплечья) указывают, что передняя конечность была способна к мощному схватыванию. Кисть передней конечности имела 5 пальцев. Первый палец был большим, заканчивающимся резко искривленным когтем и мог поворачиваться внутрь при сгибании. Второй палец был самым длинным, несколько длиннее, чем третий. Оба эти пальца имели когти, в то время как четвертый и пятый пальцы могли когтей не иметь, были очень маленькими и сравнительно простыми. Запястье окостеневало более полно, чем у других орнитисхий. В кисти присутствует мелкая гороховидная кость, которая отсутствует или не окостеневает у других архозавроморф.
В задних конечностях большеберцовая кость была на 30% длиннее бедра, что рассматривается как адаптация к скоростному передвижению. Большеберцовая и малоберцовая кости голени срастались в тибиофибулу и присоединялись к таранной и пяточной костям лодыжки (голеностопного сустава), образуя тибиотарзус, конвергентно сходный с соответствующей конструкцией у современных птиц. Также, аналогично птицам, кости нижней предплюсны (лодыжки) и плюсны срастались, формируя плюсно-предплюсну или цевку (tarsometatarsus). Задняя конечность располагалась не совсем вертикально, как и у современных птиц, бедренная кость была немного отведена в сторону. Колено заметно согнуто. Стопа задней ноги имеет 4 пальца, причем опорными являются только второй, третий и четвертый. Пятый палец утрачен. Первый короче всех остальных, фаланги несколько расходятся в стороны.

Heterodontosaurus tucki. Взрослые и молодые особи демонстрируют всеядное питание
Растительноядность также подтверждается ростральной областью предчелюстной кости, которая является беззубой и несла роговой клюв, служащий для подрезания растительности. Хотя щечные зубы гетеродонтозаврид явно приспособлены к размалыванию жесткого растительного материала, их диета, возможно, была факультативно всеядной, о чем свидетельствуют заостренные зубы предчелюстной кости и острые, искривленные когти передних конечностей. Длинные мощные передние конечности могли использоваться для разрывания муравьиных гнезд, а также для выкапывания корней и клубней. Питались эти ящеры в нижнем уровне подлеска.
Окостеневшие сухожилия соединяют невральные шипы нескольких последних спинных позвонков, но не простираются на хвостовые позвонки. Очень длинный хвост, таким образом, в отличие от многих других птицетазовых динозавров, не имел окостеневших сухожилий, обеспечивающих жесткость и, очевидно, был гибким.
У Heterodontosauridae полностью сформирован четырехлучевой тип таза с отогнутой назад лобковой костью. Таз похож на известный у фаброзаврид, но запирательный отросток отсутствует.
По крайней мере, один из представителей семейства (Tianyulong) мог иметь волокнистый наружный покров, предположительно из прото-перьев.

Heterodontosaurus tucki
Следующие особенности, могут являться синапоморфиями гетеродонтозаврид в филогенетическом контексте:
Следующие аутапоморфии характеризуют гетеродонтозаврид:
Взаимоотношения родства в семействе гетеродонтозаврид
Кладистический анализ показывает, что гетеродонтозавриды являются монофилетической группой, сестринской к Genasauria, то есть базальной ко всем остальным известным орнитисхиям, кроме писанозавра, что указывает на их очень древнее происхождение.
В самом семействе Heterodontosauridae базальные и более поздние гетеродонтозавриды, обитавшие уже только в Лавразии, становились менее специализированными, с низкими зубными коронками. Это странная тенденция, не наблюдавшаяся прежде у травоядных групп динозавров. Она была вызвана, вероятно, изменением фаунальных сообществ и широким распространением травоядных, которые могли более эффективно получать пищу. К этим динозаврам относятся Echinodon, Fruitadens, Tianyulong, Geranosaurus и неописанный ящер из формации Кайента, которые все обитали в Лавразии. Более специализированные динозавры с высокими коронками зубов и лучше приспособленные к питанию растительностью, существовавшие в ранней и средней юре Гондваны, выделяются в подсемейство Heterodontosaurinae и включают Abricosaurus, Lycorhinus, Lanasaurus, Heterodontosaurus, Manidens и Pegomastax.

Места находок гетеродонтозарид в Южной Африке и Лесото. Местность и таксон:
Nosi - Abrictosaurus consors;
Mlamli, Tushielaw, Tyinindini - Heterodontosaurus tucki;
Bamboeskloof Farm, Buck Camp, Paballong - Lycorhinus angustidens;
Maboloka - Heterodontosauridae incertae sedis (Lycorhinus sp);
Voyizane - Pegomastax africanus
Большинство ископаемых остатков гетеродонтозаврид найдены в геологических отложениях, оставленных полузасушливой окружающей средой, включая формации Кларенс и Верхняя Эллиот (серия Штормберг) Южной Африки и слои Пурбек Южной Англии. Эти породы представляют собой переменную последовательность отложений заливных лугов и равнин, а также эоловых песчаников (отлагаемых ветрами) с незначительным вкраплением речных песчаников. Это свидетельствует о циклических изменениях влажного и сухого климата в течение ранней юры. Породы серии Штормберг — сильно окисленные красные слои, которые сохраняют мало органического материала, а тот материал, что имеется, обычно неполный или фрагментарный. Однако даже в этом случае в этой местности реконструируется весьма богатая фауна. Кроме гетеродонтозаврид здесь были ранние млекопитающие, цинодонты, сфенозухии, крокодилы и прозауроподы.

Echinodon becklesii
 |
 |
Echinodon becklesii
+ Echinodon becklesii. Эхинодон Беклеса. “Колючий зуб; видовое имя в честь первооткрывателя Samuel H. Beckles”. Ранний мел (Berriasian) 150–144 млн. лет назад, Южная Англия (Дорсет). Единственный гетеродонтозаврид из Европы. Название указывает на почти шипообразный вид больших основных зазубренностей оконечной части зуба.
Длина Echinodon becklesii достигала 60–70 см, высота 30 см. Длина черепа 62 мм. Был травоядным двуногим, быстро бегавшим животным, но сохранял возможность опускаться на все четыре лапы при кормлении. Хвост был длиннее остального тела.

Литография, показывающая зубы и фрагменты челюсти, относящиеся к Echinodon becclesii, найденные в Дорсете Сэмюелем Беклесом.
Рис. 1. Передняя часть верхней челюсти и зубы.
Рис. 2. Задняя часть верхней челюсти и зубы.
Рис. 3. Часть верхней челюсти и зубы, внешняя сторона.
Рис. 4. Внутренняя сторона.
Рис. 5. Часть нижней челюсти; a, b, c, - передние зубы.
Рис. 6. Часть нижней челюсти, внешняя сторона.
Рис. 7. Граница зубов; a, b, кроны поднимающихся зубов.
Рис. 8. Внутренняя сторона.
Рис. 9. Передняя часть нижней челюсти; a, зуб.
Первоначально эхинодон рассматривался как базальный тиреофор из-за того, что при раскопках материалу эхинодона были ошибочно приписаны найденные неподалеку остеодермы (кожные кости) конечностей солемидовой черепахи, которые приняли за его кожную чешую. В действительности Echinodon не имел кожных щитков и не является родственным ни Scutellosaurus, ни семейству Scelidosauridae.

Echinodon becklesii
Известен по трем очень неполным фрагментарным черепам, в основном представляющим собой переднюю часть морды (обе челюсти, зубы, слезная и скуловая кости, небо, птеригоид и т.п.). Всего более 10 находок. Известны находки изолированных зубов эхинодона из нескольких средне- и позднеюрских локаций в Англии (Оксфордшир, Уилтшир), однако, чтобы подтвердить их идентификацию, необходимы дополнительные исследования. Неоднократные попытки обнаружить в месте его первоначальной находки дополнительный материал, к сожалению, не принесли результатов.

Место находки Echinodon becklesii на южном берегу Англии
Описан Оуэном еще в 1861 году. Найденный самым первым среди всех гетеродонтозаврид, Echinodon becklesii на самом деле представляет собой последнюю, самую позднюю, дожившую до раннего мела эволюционную линию семейства Heterodontosauridae. Найден в формации Известняки Пурбек (слои Средний Пурбек). Из этой же формации описан и знаменитый мегалозавр.
 |
 |
| Echinodon becklesii. Левый дентарий. Mс - меккелев канал | Штриховка показывает поврежденную кость, пунктирные линнии - предполагаемые края; серый тон указывает матрицу. ad2, 3 - альвеолы зубов дентария 2, 3; adf - переднее отверстие дентария; apd - сочленовная поверхность предентария; be - щечная выемка; d4 - зуб дентария 4; vc - сосудистый канал |
Левая и правая части предчелюстной кости сохранились во взаимном сочленении. Имеется переднее отверстие предчелюстной кости. Носовая ямка имеет вид широкого углубления. Оба этих элемента подобны таковым у Heterodontosaurus, но отличаются от их конфигурации у других птицетазовых динозавров, таких, как Hypsilophodon. Первый зуб предчелюстной кости расположен на расстоянии, равном двум или трем альвеолам от ее переднего края, в отличие от фаброзавра, у которого этот интервал равен только одной альвеоле.

Реконструкция челюстей Echinodon becklesii (Sereno, 2012).
adf- передняя ямка дентария; adi - дугообразная диастема; apmf- передняя предчелюстная ямка; be -щечная выемка; cp- выступ короноида; d - дентарий; d1, 2, 11 - зубы дентария 1, 2, 11; fo - ямка; m - верхняя челюсть; m1, 2, 8 - верхнечелюстные зубы 1, 2, 8; pd - предентарий; pm - предчелюстная кость; pm1, 3 - зубы предчелюстной кости 1, 3.
Щечная выемка присутствуют по всей длине зубного ряда верхней челюсти. На ней располагается немного изогнутый ряд нервно-сосудистых отверстий. Имеется диастема, в которую входил нижний клык. Хотя предентарий не сохранился у эхинодона, его наличие обозначено характерными особенностями на переднем краю дентария. Большое переднее отверстие дентария открывается вперед и переходит в отпечаток сосуда, который обеспечивал кровоснабжение предентария. В целом предентарий имел довольно свободное сочленение с зубной костью. Дентарий у Echinodon имеет особенно крепкие пропорции, даже по сравнению с другими гетеродонтозавридами. Длина дентария 28 мм.

Зубы Echinodon becklesii. A - правая верхняя челюсть; B - левый дентарий
Зубы были относительно крупными, два из них были длинными и острыми клыками. Echinodon имел 2 верхних клыка, расположенных на верхнечелюстной, а не на предчелюстной кости. Зубы предчелюстной кости (3 штуки) имеют несколько раздутые кроны и гладкую поверхность без зубчиков или зазубренностей.
 |
 |
| Реконструкция Echinodon becklesii | Nuthetes destructor, гипотетически реконструированный как дромеозавр, атакует эхинодона |
Верхнечелюстная кость имеет 9 зубов. Первый из них — это почти прямой, поперечно сжатый клыкообразный зуб, без какой-либо орнаментации на мезиальном или дистальном киле. Он имеет относительно прямую и стройную крону и заметно меньшие размеры, чем нижний клык. Следующие за ним зубы имеют по 8–10 зубчиков на каждой стороне, но они занимают только верхнюю четверть кроны.

Дентарий Echinodon becklesii. Прорисовка левого дентария. Вид сбоку (A), середины (B) и сверху (C) (NHMUK 48215b). Штриховка показывает поврежденную кость, пунктирные линнии - предполагаемые края; серый тон указывает матрицу.
ad1, 2, 10 - альвеолы зубов дентария 1, 2, 10; adf - переднее отверстие дентария; apd - сочленовная поверхность предентария; be - щечная выемка; d4, 9 - зубы дентария 4, 9; io - отпечатанная орнаментация; Mc - Меккелев канал; ssymf - кромка симфизиса; sym - симфизис; symt - впадина симфизиса
Дентарий содержит 11 зубов. Первый зуб был маленьким и колышкообразным. Ему предшествует существенный беззубый участок, что отличает эхинодона от других гетеродонтозаврид и других базовых орнитисхий. Следующий зуб является большим и клыкообразным. Кроны постклыковых зубов дентария хорошо отделены от их корней и имеют большую высоту, чем соответствующие им верхнечелюстные зубы. Верхняя половина этих крон имеет зазубрины. Строение зубов у Echinodon becklesii указывает на растительноядную диету, но низкие коронки щечных зубов и отсутствие граней износа говорят об определенной всеядности эхинодона.
других гетеродонтозаврид и других базовых орнитисхий. Следующий зуб является большим и клыкообразным. Кроны постклыковых зубов дентария хорошо отделены от их корней и имеют большую высоту, чем соответствующие им верхнечелюстные зубы. Верхняя половина этих крон имеет зазубрины. Строение зубов у Echinodon becklesii указывает на растительноядную диету, но низкие коронки щечных зубов и отсутствие граней износа говорят об определенной всеядности эхинодона.
Синоним: Echinodon becklessii, Echinosaurus, Saurechinodon.

Fruitadens haagarorum
 |
 |
Fruitadens haagarorum
+ Fruitadens haagarorum. Фруитаденс Хаагов. «Зуб из города Фруита; видовое имя в честь целого ряда сотрудников по фамилии Хаага из Музея естественной истории в Лос-Анджелесе». Поздняя юра (ранний–средний Tithonian), Северная Америка (Колорадо). Двуногий мелкий орнитисхий. Длина взрослой особи 65–75 см, вес — около 1 килограмма. Длина черепа молодого экземпляра — 60 мм, взрослого экземпляра — 75 мм. Первый гетеродонтозаврид из Северной Америки, описан Батлером в 2010 г.
 |
 |

Размеры Fruitadens haagarorum
Фруитаденс — один из самых маленьких птицетазовых динозавров, благополучно обитавший в тени юрских гигантов. До открытия Fruitadens, большинство палеонтологов не представляли себе, что динозавры могли быть такими крошечными. Особенно это касалось птицетазовых ящеров, для которых крупные размеры считались обусловленными растительноядным типом питания, хотя это никак не мешает существованию мелких растительноядных и всеядных млекопитающих и современных рептилий. Исследование сросшихся длинных костей, а также общие пропорции говорят, что окаменелости фруитаденса принадлежат взрослому животному.
  |
 |
Fruitadens haagarorum
Его диета, возможно, была всеядной. Помимо изучения строения зубов, была предпринята проба восстановить челюстные мышцы и определить характер их действия. Fruitadens был способен широко раскрывать свою челюсть и производить быстрый укус в отличие от его более старшего родственника гетеродонтозавра, который имел более мощный прикус, но меньшее раскрытие челюстей. Более слабый, но быстрый укус возможно свидетельствовал об охоте на беспозвоночных наряду с питанием растительностью. Это подтверждает и отсутствие хорошо развитых граней износа на зубах. В целом приспособление зубов к растительной диете несколько ниже, чем у гетеродонтозавра.
|
|
|
Сохранившиеся элементы Fruitadens haagarorum
Fruitadens haagarorum известен из четырех отдельных мест по частичным остаткам черепов и скелетов, по меньшей мере, от 6 особей на различной стадии роста.
 |
 |
 |
| Fruitadens haagarorum. Верхняя челюсть вид сбоку. asc - сломанное основание восходящего выроста; dia - диастема; for - отверстия; mxsh - челюстная полка |
Дентарий, медиальный вид for - отверстие; mgr - Мекелиева борозда; rfor - отверстие замещения |
a - левая верхняя челюсть; c - правый дентарий. dia - часть диастемы; с - клык. Челюсть - голубая, дентарий - фиолетовый, фрагмент неба - зеленый, функциональные зубы - желтые, замещающие зубы - оранжевые, внутренние каналы - красные |
Его челюсти были менее специализированы, чем у раннеюрского Heterodontosaurus. Морфологией зубов и челюстей он ближе к Tianyulong, чем к раннеюрским гетеродонтозавридам. Имеется большая и глубокая предглазничная ямка, слабая щечная выемка. На каждой ветви дентария имелось по 2 нижних клыка; верхние клыки, в отличие от эхинодона, похоже, отсутствовали. Имеются активно замещаемые зубы, в отличие от многих гетеродонтозаврид. Перед клыком находился маленький колышкообразный зуб, который отсутствует у всех других орнитисхий. Клык не имеет зазубрин и по диаметру он не превосходит самые большие другие зубы. Зубы предчелюстной кости (3 штуки) относительно небольшие, без увеличения в размерах в направлении спереди-назад. Верхняя челюсть содержала 7–8 зубов, кроны которых отличались от таковых у гетеродонтозавра и больше напоминали зубы базального орнитисхия фаброзавра. На дентарии располагается 11 зубов. В середине дентария располагались самые большие его зубы, они постепенно уменьшались дистально и проксимально. Боковая поверхность дентария пронизана многочисленным питательными отверстиями, расположенными через нерегулярные интервалы. В целом дентарий сильно напоминает таковой у Echinodon.
 |
 |
| Fruitadens haagarorum. a - Правая предчелюстная кость; b, c - Левая верхняя челюсть; d, e - Правый дентарий; f - Задние зубы дентария; g, h - Проксимальная часть правого бедра; i - Дистальный конец левой большеберцовой кости; j - Левое бедро; k–n - Левая большеберцовая кость, малоберцовая кость, таранная и пяточная кости; o, p - Дистальный конец левой большеберцовой кости, малоберцовой, таранной и пяточная костей. Сокращения: ac.os - восходящий вырост таранной кости; atr - передний вертел; c - клык; dia - часть дугообразной диастемы; fo - отверстие; ft - сломанное основание четвертого вертела; mf - кромка |
Прорисовка костей задних ног. amsh - передне-средняя пластина большеберцовой кости; ast - таранная кость; cal - пяточная кость; cnc- коленный гребень; dasp - дорсальная часть восходящего выроста, сформированная отдельным окостенением; fib - малоберцовая кость; fib.a - сочленовная поверхность для малоберцовой кости; fibc - мыщелок малоберцовой кости; for - отверстие; innc - внутренний мыщелок |
Фруитаденс близок гетеродонтозавру общими пропорциями тела и анатомически, с относительно короткими передними лапами и длинными задними ногами. Кости задних ног были полыми, как у мелких теропод. Бедро имеет стержнеподобный четвертый вертел, который присутствует только у гетеродонтозаврид и отсутствует у других птицетазовых динозавров. Таранная и пяточная кости сросшиеся друг с другом. Их соединение с большеберцовой и малоберцовой костями имеет ясно различимую линию шва, то есть они еще срослись не полностью.
 |
 |
 |
Fruitadens haagarorum

Передний хвостовой позвонок Fruitadens haagarorum.
poz - постзигапофизис; prz - презигапофизис; tvp - поперечный отросток
Имеется 6 крестцовых позвонков с массивными крестцовыми ребрами. Шейные и спинные позвонки в целом похожи на таковые у других небольших орнитисхий.

Место находки окаменелостей Fruitadens haagarorum
Местность обитания Fruitadens haagarorum (геологическая формация Моррисон — Палеонтологическая область Фруита) представляла собой заливные луга и равнины с извилистыми руслами рек, берега которых были покрыты лесами. В то же время, вдали от рек имелись временные водоемы, земли были относительно сухими, почти как в наше время. Их населяли наземные крокодилы-мезозухии и сфенозухии, миниатюрные млекопитающие, ринхоцефалы Eilenodon и Opisthias, различные ящерицы и крошечные динозавры. Разрозненные кости других динозавров обычны в этой местности — тероподы Ceratosaurus и Allosaurus, зауроподы Camarasaurus и Apatosaurus, орнитисхии Stegosaurus и Dryosaurus. Водоемы населяли черепахи, двоякодышащие рыбы и актиноптеригии.

Fruitadens haagarorum
Fruitadens haagarorum характеризуется следующей уникальной комбинацией особенностей:

Tianyulong confuciusi
 |
 |
Tianyulong confuciusi
+ Tianyulong confuciusi. Тьенюлунг Конфуция, Тянюлонг или Тяньюйлун. «Дракон Тьеню. Назван в честь Музея Естественной Истории Шаньдун Тьеню (Shandong Tianyu); видовое имя в честь Конфуция». Поздняя средняя юра (Callovian), Китай (Западный Ляонин). Мелкий двуногий птицетазовый динозавр, первый гетеродонтозаврид, найденный в Азии. Длина 70 см. Найденный экземпляр является взрослой особью, что доказывается слиянием крестцовых позвонков, слиянием или плотным сочленением невральной дуги и всех других сохранившихся позвонков, и слиянием между большеберцовой костью и ближайшими предплюснами и между основами плюсен. Геологический возраст Тьенюлунга долго считался неуверенным, так как он был найден в приватном порядке и содержащая порода могла быть как раннемелового (Barremian–Aptian), так и юрского возраста. Последний вариант подтвердился при радиоизотопном датировании.
 |
 |
Реконструкция скелета Tianyulong confuciusi. Показаны сохранившиеся кости и покровные волокна. Позвонки конца хвоста неизвестны
Тьенюлунг питался растительностью, факультативно был всеядным. Имел стройное тело, длинный хвост и клыкообразные зубы. Пропорции скелета несколько необычные: относительно большой череп, короткие передние конечности и длинный палец II руки.

Tianyulong confuciusi. Прорисовка найденных волокон
Сохранился волокнистый наружный покров, который интерпретируется как протоперья, аналогичные найденным у некоторых теропод. Эти неразветвленные волокна образуют гребень, идущий вдоль шеи, спины и хвоста. На спине их длина была 3,8 см, в районе хвоста — 5 см. Волокнистые покровные структуры сохранились в трех участках на окаменелости: один несколько ниже шеи, другой на спине и наибольший выше хвоста. Полые волокна расположены параллельно друг другу. Они выглядят относительно жесткими (имеется небольшое искривление, но нет никаких изгибов или волнистостей), что делает их аналогичными покровным структурам, найденным на хвосте пситтакозавра, чем протоперьям, обнаруженным у теропод. Среди теропод покровы Tianyulong ближе всего к неразветвлённым протоперьям у Sinosauropteryx и Beipiaosaurus. От покровов синозавроптерикса они отличаются значительно и размерами, и плотностью. У этого теропода они более широкие, более длинные и реже расположенные.

Волокнистые образования, найденные на матрице с Tianyulong
Предполагаемая длина некоторых покровных структур на хвосте – около 60–70 мм, что в семь раз больше высоты хвостового позвонка и составляет десятую часть длины всего тела. Диаметр около 0,5 мм. Их длина и полая природа указывают, что они развились из подкожных структур, таких, как волокна коллагена.
 |
 |
Tianyulong confuciusi
Прежде кожные покровные структуры были известны только у развитых теропод и птицетазовых, поэтому их открытие у Tianyulong продляет их существование вниз по филогенетическому дереву. Однако гомология между волокнами орнитисхий и протоперьями теропод не очевидна. Если бы эта гомология нашла подтверждение, отсюда следовало бы, что общий предок птицетазовых и ящеротазовых динозавров был покрыт пероподобными структурами и что те группы динозавров, у которых известны отпечатки кожи (зауроподы) оказались бы вторично лишенными перьев. Однако у самых ранних видов тероподов не найдены свидетельства протоперьев, очевидно, что и у общего предка их не было.
Если гомология не находит поддержки, то это значит, что волокнистые кожные структуры развивались независимо у орнитисхий и заурисхий, так же, как и у других архозавров, таких как птерозавры. Наличие волокнистого покрова у теропода Beipiaosaurus поддерживает идею о гомологичности волокон Tianyulong протоперьям. Волокна у Tianyulong и у Beipiaosaurus были длинными, одиночными и неразветвленными (в отличие от перьев). У Beipiaosaurus, однако, они были уплощенными. У Tianyulong волокна были округлыми в поперечном сечении и поэтому находились ближе по структуре к самым ранним формам перьев, предсказанным экспериментальными моделями.

Четыре интерпретации внешнего вида Tianyulong confuciusi:
A: Только волокна, найденные у голотипа. Можно только утверждать , что Tianyulong confuciusi имел кератиновые структуры в этих местах.
B: Минимальная интерпретация расположения волокон. Наиболее вероятная, потому что подобна распространению возможных волокон, сохранившихся у другого динозавра, Psittacosaurus (экземпляр SMF R 4970). Отсутствие непрерывности спинных и хвостовых волокон не похоже на структуру, которую бы ожидали увидеть у животного.
C: Покрытый пухом, как в большинстве интерпретаций. Маловероятно, так как пух (короткие волокна, как у Sinosauropteryx prima) никогда не находили у орнитисхий, и даже волокна отмеченые у теропод и орнитисхий могут быть аналогичными структурами.
D: Наибольшее покрытие тела длинными волокнами
На самом деле не стоит спешить с утверждениями, что многие динозавры имели покровы. Сохранившиеся отпечатки кожи от гадрозавров, цератопсов, стегозавров, анкилозавров, зауропод (включая эмбрионы) и большинство теропод — аллозавры, тираннозавры — имели чешуйчатую кожу. Протоперья были исключением, а не правилом. Создание реконструкций почти всех гетеродонтид покрытых волокнам — это явное проявление фантазии, не подкрепленное реальными находками. Тем более что и характер покрова у самого Tianyulong вызывает некоторые сомнения в их происхождении.
 |
 |
| Tianyulong confuciusi и Psittacosaurus | Сравнение "щетин" Psittacosaurus (вверху), протоперьев Sinosauropteryx (слева) и "волокон" Tianyulong (справа) |
Щетина пситтакозавра ясно расположена в пределах кожи, в то время как волокна тьенюлунга не касаются скелета вообще. Некоторые волокна найдены возле спины, другие ниже шеи. Однако эти волокна очень удалены от тела. Дорсальная часть не следует за дугой позвонков, вместо этого она лежит впереди костей. Что касается вентрального участка, то здесь волокна фактически не связаны с костями и при этом они не следуют за контуром тела. Хвостовые волокна также странно расположены. Как и вся остальная часть этих волокон, они не следуют за контуром тела (сравните, например, как протоперья синозавроптерикса строго соответствуют контуру тела). Фактически, многие из этих волокон запутаны между собой. Вместо того чтобы быть более или менее равномерно распределенными вдоль спины или хвоста животного, большая их часть находится в районе основания хвоста и половина из них весьма далеко от тела. Похоже, что все эти волокна являются остатками какого-то волокнистого растения, окаменевшими вместе с телом погибшего ящера.
 |
 |
| Частичный череп Tianyulong confuciusi из провинции Ляонин, PRC (STMN 26-3; Zheng et al. 2009). a - угловая кость; ad 9, 10 - альвеолы зубов дентария; 9, 10; adf - переднее отверстие дентария; antfo - предглазничная ямка; apd - сочленовная поверхность предентария; d - дентарий; d1, 2, 8 - зубы дентария 1, 2, 8; emf - внешнее верхнечелюстное отверстие; en - наружные ноздри; j - скула; l - слезная кость; m - верхняя челюсть; n - носовая кость; pd - предентарий; pf - префронтальная кость; pm - предчелюстная кость; po - заглазничная кость; q - квадратная кость; qj - квадратноскуловая кость; sa - надугловая кость |
Верхнечелюстное расположение зубов гетеродонтозаврида Tianyulong confuciusi, Китай. Правые верхнечелюстные зубы ?6-9 в виде сбоку (IVPP V17090). Штриховка показывает поврежденную кость, пунктирные линнии - предполагаемые края; серый тон указывает матрицу. cel - эктолоф (гребень) цингулюма; ci - цингулюм; m6 9 - верхнечелюстные зубы 6 9; ne - шейка; pri - первичный гребень; rt - корень |
Форма черепа у Тьенюлунга подобна таковой у Heterodontosaurus; она полутреугольная при виде сбоку и слегка вогнута в предглазничной области. Длина черепа составляет 66 мм. Приблизительно 60% длины предчелюстной кости являются беззубыми, задние 40% ее длины размещают 2 зуба, в отличие от большинства гетеродонтозаврид, имевших 3 зуба. Второй зуб предчелюстной кости клыкообразный, для него имеется диастема. Клык немного изгибается назад. Сбоку верхней челюсти расположена полутреугольная предглазничная впадина, ограниченная вентрально острым, немного изогнутым ободком. Щечная выемка более узкая, чем у Echinodon и Lycorhinus. Имеется небольшой клиновидный предентарий. Ветви дентария прямые и с параллельными сторонами на большей части своей длины, в отличие от большинства гетеродонтозаврид, которые демонстрируют углубление ветви сзади. Имеется внешнечелюстное отверстие.
 |
 |
| Передняя часть черепа гетеродонтозаврида Tianyulong confuciusi (IVPP V17090), Китай в виде сбоку. Штриховка показывает поврежденную кость, пунктирные линнии - предполагаемые края; серый тон указывает матрицу. ad1 - альвеолы зубов дентария 1; antfe - предглазничное отверстие; be - щечная выемка; d - дентарий; d1, 5, 9 - зубы дентария 1, 5, 9; en - наружные ноздри; fo - ямка; l - левый; m - верхняя челюсть; m4, 9 - верхнечелюстные зубы 4, 9; n - носовая кость; nf - назальная ямка; pd - предентарий; pm - предчелюстная кость; pm1, 2 - предчелюстные зубы 1, 2; r - правый |
|
Дентарий Tianyulong confuciusi имеет примерно 10 зубов. Клыкообразный зуб является первым в ряду, никаких следов предшествующего маленького зуба не имеется. Следующий зуб расположен без промежутка, который имеется у Echinodon и Fruitadens. Остальные зубы дентария имеют поясок (цингулюм). Предпоследний, девятый зуб дентария является наибольшим. В верхней челюсти всего должно было быть 11 зубов. Форма их крон, как и зубов дентария, полутреугольная.
 |
 |
 |
Tianyulong confuciusi
 |
 |
Tianyulong confuciusi
В целом, пропорции тела Tianyulong очень необычны среди всех гетеродонтозаврид. Череп очень большой и задние конечности длинные, в то время как шея и туловище пропорционально маленькие, а передние конечности короткие. Такие необычные пропорции придавали этому мелкому ящеру внешний вид большеголового, короткорукого, но длинноногого карлика, с совсем другими пропорциями тела, чем у Heterodontosaurus.
В голотипе Tianyulong confuciusi сохранилась задняя половина шейных и большая часть спинных позвонков. Длина тела позвонка почти постоянна и составляет 5 мм. Имелось 9 шейных и 12 спинных позвонков. Крестцовые позвонки сросшиеся. Хвост был очень длинным по отношению к телу и превышает длину остального позвоночника на 20 позвонков. Хвостовые позвонки увеличиваются в длину на 40% от первого до десятого позвонка. Окостеневшие сухожилия имеются на задних спинных, крестцовых и передних хвостовых позвонках.
 |
 |
Предплечье и кисть руки гетеродонтозаврида Tianyulong confuciusi. Левая локтевая кость, лучевая кость и кисть руки главным образом в виде спереди или снизу. Штриховка показывает поврежденную кость, пунктирные линнии - предполагаемые края; серый тон указывает матрицу.
I-III - пальцы I-III; mc1-5 - пястные кости 1-5; ph - фаланга; ra - лучевая кость; ul - локтевая кость; un - когтевая фаланга
Длина плечевой кости 26–28 мм. Кисть руки имела все 5 пальцев. Пропорции пальцев руки являются самыми необычными среди птицетазовых динозавров, включая Heterodontosaurus. Палец и пястная кость II более длинные, чем палец и пястная кость III. Предпоследние фаланги в пальцах II и III, кроме того, более длинны, чем предыдущие фаланги, предполагая усиленную хватательную функцию кисти руки у Tianyulong, как и у Heterodontosaurus. Когтевые фаланги поперечно сжатые и острые, на первом пальце они длиннее, чем на пальцах II и III.
 |
 |
Стопа гетеродонтозаврида Tianyulong confuciusi. Левые фаланги ног (слева); правые фаланги ног (справа).
I-IV - пальцы I-IV; csh - оболочка когтя; mc1-4 - пястные кости 1-4; ph - фаланги; un - когтевая фаланга
Седалищная кость не имеет выроста запирателя, существующего у многих орнитисхий. Лобковая кость имеет постлонный вырост. Большеберцовая кость чрезвычайно вытянута, достигая более чем 140% от длины бедра. Большеберцовая, малоберцовая кость и ближайшие предплюсны сросшиеся, как тибиотарзус (голено-предплюсна) у большинства экземпляров Heterodontosaurus. Первый палец ноги очень короткий, но его когтевая фаланга весьма большая, превосходит фалангу на пальце IV и равна фаланге на пальце II. Третий палец ноги длиннее остальных, что соответствует приспособленности к бегу. Следы пятого пальца ноги отсутствуют. Когтевые фаланги поперечно сжаты и сужаются до стройных кончиков. Некоторые из них сохранили части роговой оболочки когтя.

Tianyulong confuciusi
Характерные особенности Tianyulong confuciusi:

 |
 |
Tianyulong confuciusi. 2 найденных скелета
Находка состоит из 2 неполных сочлененных скелетов с частичным черепом и челюстями и еще 4 скелетов разной полноты. Захороненный в озерных осадках и засыпанный вулканическим пеплом, Tianyulong сохранил сотни щетин, покрывавших его тело.
 |
 |
Tianyulong confuciusi

Левый дентарий неописанного гетеродонтозаврида из ранней юры формации Кайента из Аризоны в виде сбоку ( Sereno et al. unpublished)
adf - переднее отверстие дентария; apd - сочленовая поверхность для предентария
+ Kayenta. Гетеродонтозаврид из формации Кайента. Ранняя юра (Sinemurian–Pliensbachian), Северная Америка (Аризона). Очень маленький, частично сочлененный скелет птицетазового динозавра. Формация Kayenta. Идентифицирован как молодая особь гетеродонтозаврида. Сохранились верхние и нижние челюсти с зубами, много других частей черепа, позвонки от всех отделов позвоночника, части плечевого и тазового поясов, конечностей. Щечные зубы имеют низкие коронки, как у Echinodon, Fruitadens, Tianyulong. Пока не описан.
Продолжение статьи см. Гетеродонтозавриды (Heterodontosauridae, Lycorhinidae) Часть 2


